
Some of the most common species of frugivorous birds known to consume and disperse Pistacia lentiscus fruits. In clockwise order starting from top left and with their migrant behaviour code in brackets: Saxicola rubicola (R), Curruca melanocephala (R), Turdus philomelos (W), Sylvia borin (TS), Sylvia atricapilla (W), Phoenicurus phoenicurus (TS), Erithacus rubecula (W), Turdus merula (R) and Chloris chloris (R; this last species acts as seed predator). Migrant codes: R – resident, W – wintering migrant, TS – trans-Saharan migrant. Photo credits: Elena Quintero.
Measuring the natural regeneration of plants and their success can be a tricky and demanding task, especially if seeds are dispersed by a wide array of fruit-eating animals (frugivores). In our recent study we seek to disentangle the lasting effects that frugivore birds have in the early recruitment of the plant species Pistacia lentiscus (lentisc). This plant is a widely distributed shrub in the Mediterranean Basin, from Portugal to Syria and down south to Morocco. Check out the map below of Pistacia lentiscus distribution (in grey) and bird migratory routes together with a caption of P. lenticus fruits. From Martínez-López et al. 2020.
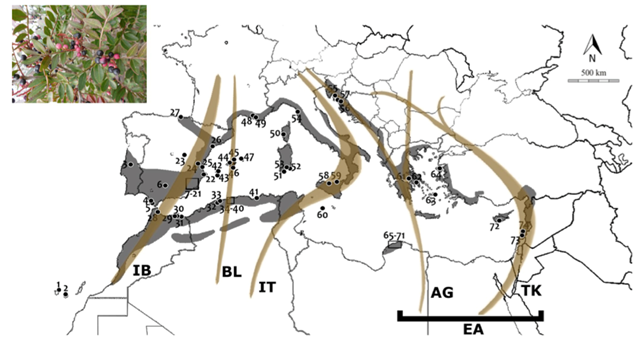
During the winter in the northern hemisphere, many birds rely on the abundant fruits that this plant produces. The fruiting season is long and can extend from September to March. The fruits are consumed not only by resident birds, but also by migrant species.
Our study site is Doñana National Park, in the Southern-west part of the Iberian Peninsula in Spain. Between the months of September and October, many trans-Saharan migrant birds (such as many warblers species and the common redstart) consume big quantities of P. lentiscus fruits during their stop-overs before crossing the Gibraltar strait to their wintering grounds in Africa. Bird species from the North of Europe (such as the shameless robin or the song thrush) also start arriving around October to overwinter in the Mediterranean scrubland. They need to gain enough energy to migrate back to their breeding grounds in spring. Fruit consumption from all these birds is crucial for the dispersal of lentisc seeds and further recruitment of seedlings.
One of the key aspects of our research is based on how birds affect plant recruitment, not only by measuring the amount of fruit they consume and seeds they disperse, but also by looking at where they deposit those seeds. Since birds do not use the landscape randomly, the seeds they disperse will be deposited in different amounts in different locations. That is, some birds will prefer to perch on pine trees while others prefer low scrubland. The microhabitats where seeds are deposited will ultimately affect their chances of survival because of different environmental and ecological conditions. For example, different deposition sites will be prone to different degrees of rodent predation pressure (they tend to avoid the open areas), as well as present different probabilities of germination and seedling survival because of humidity or sun exposure.
We got out into the field and measured the probabilities of seeds surviving at different steps of the demographic process, starting from when fruits are produced in the plant and consumed by birds, up to the survival of seedlings in different microhabitats. In the first figure of our manuscript we showed the different transitions stages we had studied and measured.
This is figure one of our manuscript showing the different transition staged we measured for lentisc demographic cycle. TP1: probability of fruit consumption, TP2: probability of seeds escaping finch predation, TP3: probability of dispersing a viable seed; TP4: probability of viable seed being dispersed to a certain microhabitat, TP5: probability of dispersed seeds escaping rodent predation, TP6: prob. of seedlings emerging from surviving seeds, TP7: probability of seedling surviving its 1st summer and TP8: probability of seedling surviving its 2nd summer. Coloured rectangles represent distinct microhabitat types that differ in arrival of seeds and recruitment probabilities.
For such task, we used a wide array of methods and experiments. To characterise how much different birds were consuming at focal plants, we used Go-Pro monitoring cameras. In one field season alone we collected 760 hours of video containing avian visits, that amounts to a month of non-stop video-footage! But, thanks to invaluable help from Antonio Orta, and an intelligent software to detect movement (DeepMeerkat by Weinstein 2018), we processed all the videos and managed to capture nearly 4000 plant-animal frugivory interactions.
Additionally, to characterise even more avian visits to plants and their heterogenous seed deposition in different microhabitats we collected bird faecal samples by placing trays (a.k.a. seed-traps) underneath plants in different microhabitats. We then used DNA-barcoding, a wonderful and almost magical molecular technique that allows the identification of the animal’s identity from any fecal sample or regurgitated seed collected (see Gonzalez-Varo et al. 2014). We placed more than a hundred seed-traps around our study sites to capture dispersed seeds and processed them later in the lab. This meant careful sample collection in the field in order not to contaminate them with human DNA, along with protocols in a sterile lab and many PCRs to amplify avian DNA and sequencing. This hard work was led by Juan Miguel Arroyo, co-author in the publication.

This project was part of my PhD thesis and these are photos of some of the wonderful people who contributed collecting data the field. From left to right and up to down: Pedro Jordano (co-author in the study), Gemma Calvo, me (Elena Quintero) Miguel Jácome; Irene Mendoza, me, Paco Rodríguez-Sánchez (co-author in the study); Jorge Isla, me again sorry!, Antonio Orta and Blanca Arroyo-Correa.
Lastly to further follow the seeds fate after dispersal, we performed two different experiments in the field. First, to estimate seed predation pressure, we did some seed offerings and checked them regularly to detect the pace and proportion at which seeds were predated by rodents. Then, to estimate the probabilities of seedling emergence and survival we did seed sowing experiments in different microhabitats and once seedlings emerged, we followed their survival up to their second summer. Summer months are the ones with highest mortality for Mediterranean seedlings due to extreme heat and arid conditions. We then merged all of this data together, and, using a Bayesian approach, managed to estimate the approximate contribution of different birds in the different stages along the demographic cycle of lentiscs (see figure 5A for results in our published manuscript).
So, all in all, after months of fieldwork, many hours in the lab, video screening time and several additional hours of analysis, we estimated that a lentisc plant would need to produce around 500,000 fruits in order to recruit a 2-year old seedling (with an 80% credibility interval ranging from 300,000 to a million fruits). This is way more fruits than a plant normally produces in our study site, where individual plants produce between 3,000 to 120,000 fruits each season.
Studying the effect of birds and microhabitat suitability on plants’ recruitment will be useful to better understand natural regeneration in the face of future fluctuations in animal abundances, changes in landscape-use and anthropogenic disturbances scenarios, given that we know recruitment success is dependent on the bird community and the distribution of available microhabitats in the landscape.

Our field experiments. From left to right: Go-Pro camera recoding a lentisc plant, close-up of faeces with P. lentiscus seed collected in a seed trap, a seed offering rodent predation experiment, a 1 year old seedling of P. lentiscus grown in one of our seed sowing experiments. Photo credits: Elena Quintero.
You can read more about the lasting effects of avian‐frugivore interactions on seed dispersal and seedling establishment by Quintero, E., Arroyo, J. M., Dirzo, R., Jordano, P., & Rodríguez, F. (2024) in the Journal of Ecology, pp. 1–17: https://doi.org/10.1111/1365-2745.14260 Also you can find out more about Elena’s work on the Pedro Jordano Lab website here: http://ebd10.ebd.csic.es/
References:
Martínez‐López, V., García, C., Zapata, V., Robledano, F., & De la Rúa, P. (2020). Intercontinental long‐distance seed dispersal across the Mediterranean Basin explains population genetic structure of a bird‐dispersed shrub. Molecular Ecology, 29(8), 1408–1420. https://doi.org/10.1111/mec.15413
Weinstein, B. G. (2018). Scene‐specific convolutional neural networks for video‐based biodiversity detection. Methods in Ecology and Evolution, 9(6), 1435–1441. https://doi.org/10.1111/2041-210X.13011
González-Varo, J. P., Arroyo, J. M., & Jordano, P. (2014). Who dispersed the seeds? The use of DNA barcoding in frugivory and seed dispersal studies. Methods in Ecology and Evolution, 5(8), 806–814. https://doi.org/10.1111/2041-210X.12212

