Matsuo Itoh discusses his article: ‘Parapatric and sympatric adaptation of Setaria viridis populations in Japan to heterogeneous coastal habitats via trait divergence of plant form, salt spray tolerance and flowering time.’
Maritime ecotypes
Maritime plants contain dwarf ecotypes that are adapted for exposure to strong winds and salt spray. The compact form of coastal plants is caused by the inhibition of growth and leaf necrosis induced by salt spray. Salt spray tolerance also plays a role in adaptations to the coastal environment. Ecotypes are widespread groups that evolve from geographically isolated populations within species and are often regarded as the precursors of new species. When variants differentiated from local populations are well-adapted and reproductively isolated, they become forerunners of new ecotypes. Setaria viridis, which is a self-fertilising annual grass distributed globally, comprises three variants that differ in plant form and salt spray tolerance: short and tolerant, on rocky cliffs and pebble shores of open seas; tall and mid-tolerant, on the sandy shores of the Seto Inland Sea; and ordinary inland tall and susceptible variants. The short and tolerant variant is distributed in the coastal habitats of East Asia, i.e. throughout Japan and South China, and is equivalent to the taxonomic ecotype. Coastal S. viridis populations are suitable for studying population divergence because they show high variability that has generated multiple coastal variants.

Our study
Our study aimed to determine how coastal variants arise from local populations under natural selection by studying phenotypic variations and survival of S. viridis populations inhabiting pebble and sandy seashores in Japan. To identify adaptive traits characterising variants, common garden experiments were performed for determining the variation in morphology, salt spray tolerance, and flowering time among populations in four adjacent habitats at two localities. Studies of populations in these adjacent habitats are favourable to estimate selective agents within the specific habitats as differences in some environmental factors, such as temperature and humidity, can be neglected in small spatial areas. To determine selective agents and their adaptation, field transplant experiments were performed, population dynamics and flowering phenology were monitored, and artificial crossing was conducted for second generation segregation.
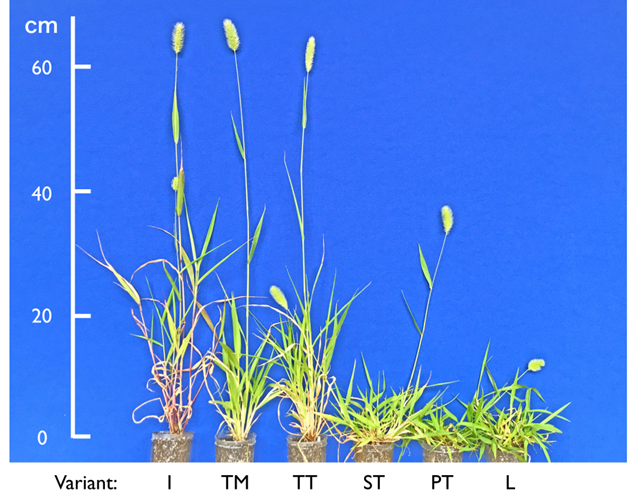
S. viridis populations comprised five coastal variants showing significantly higher salt spray tolerance than the inland variant (I variant in the photographs). The most common of these was a short and tolerant variant (ST) adapted to cliff and pebble shore under intensive salt spray from the Pacific Ocean. Three others were a tall and mid-tolerant variant (TM) that endured mild breezes off the Seto Inland Sea, a tall and tolerant variant (TT) that survived sea splash from sporadic storms, and a prostrate and tolerant variant (PT) specialised for rock cracks exposed to strong sea winds.
When plants were transplanted and exposed to sea winds on the cliff top, only the I variant was fatally damaged owing to the lack of salt spray tolerance. The tall and mid-tolerant TM plants were eliminated by sea splash from typhoon, whereas the TT variant survived owing to its high salt spray tolerance. The TT variant possessed a height disadvantage over the ST variant in open habitats under intensive salt spray from open seas. These variants imply that maritime plants first acquired salt spray tolerance for survival, after which compact plants evolved in habitats where strong winds caused damage from salt spray. Flowering in a fifth variant (L) was delayed, avoiding the fatal risk of reproducing during the mid-summer drought. Although salt spray tolerance and late-flowering were partially recessive, self-fertilisation of S. viridis may have increased the homozygosity of recessive genes to generate novel variants.
Implications for ecotype formation
Diverse intensities of salt spray and wind as well as summer drought have been generating various coastal variants in parapatry and sympatry. This was partially attributable to self-fertilization which may rapidly increase the number of homozygotes owing to diminishing gene flow between-populations. Among these, a compact variant adapted to common local seashores may become the forerunner of geographically widespread ecotypes. Other coastal variants would remain as local populations or be eradicated due to occasional environmental changes. Local adaptation in mosaic habitats will assist in determining adaptation modes of plants and the beginning of ecotype formation.


