Beatriz Aguirre, Cornell University, discusses her article: Plant functional diversity does not dilute virus infection, but community virus prevalence positively correlates with net C3 grass production
Greater plant diversity can protect plant communities from pathogens in wild and agricultural plant communities, and the effects of many aspects of diversity, such as genetic and species diversity, have been studied. However, diversity among plant functional groups (i.e. grasses, legumes, and forbs) within communities can also shape pathogen dynamics by altering host density, soil nitrogen pools, and the number of encounters between host plants and the insect vectors that transmit plant pathogens. While plant functional diversity is well documented to positively influence key ecosystem functions, specifically plant productivity, we still don’t know how plant functional diversity influences plant virus dynamics. Additionally, insect-vectored virus infection dynamics may differ among C3 and C4 hosts due to differences in their physiological responses to environmental factors and differences in their leaf nitrogen content, which may directly influence host palatability to vectors.
To examine the effects of plant functional diversity on plant virus prevalence, and how plant virus prevalence affects community and host production, we established an experimental field study in Freeville, New York. We set up experimental plant communities with varying levels of plant functional diversity that included either C3 or C4 grasses for two years. We covered plots using a fine mesh cage upon plant emergence, to prevent exogeneous pathogens and insects from entering the communities. We then inoculated the communities with Rhopalosiphum padi aphids carrying a widespread grass virus, barley yellow dwarf virus (BYDV-PAV).
The experimental set-up at the Homer C. Thompson Research Farm in Freeville, New York (top). A community with just C4 grasses (bottom left). A community with C4 grasses and forbs (bottom right). Photos by Beatriz A. Aguirre.
At the end of each growing season, we assessed plant virus infection in each community, as well as the aboveground production of each plant species. We evaluated net community production and the production of the plant functional groups in each year of the study.


Sampling grasses in the field (left) and testing grass samples from the field for barley yellow dwarf virus infection using Double Antibody Sandwich Enzyme Linked Immunosorbent Assay (DAS-ELISA) in the lab (right). Photos by Alan S. Hinojosa.
We found that, despite barley yellow dwarf virus (BYDV-PAV) infection levels varying significantly among study years, neither plant functional composition nor richness mitigated or amplified virus infection consistently across both study years. We found that species varied in their overall virus infection levels and net production. We also found that there were positive, neutral, and negative relationships between community-level virus prevalence and species’ net production. These findings reflect varying virus susceptibility and competence among hosts. Surprisingly, at the functional group level, we found that there was a strong positive correlation between community-level virus prevalence and net C3 grass production, in contrast to a neutral relationship between community-level virus prevalence and C4 grass production. Further, we found that net grass production was not correlated with community-level BYDV-PAV prevalence, suggesting that grass production likely contributed to variation in virus prevalence but likely did not drive community-level virus prevalence.
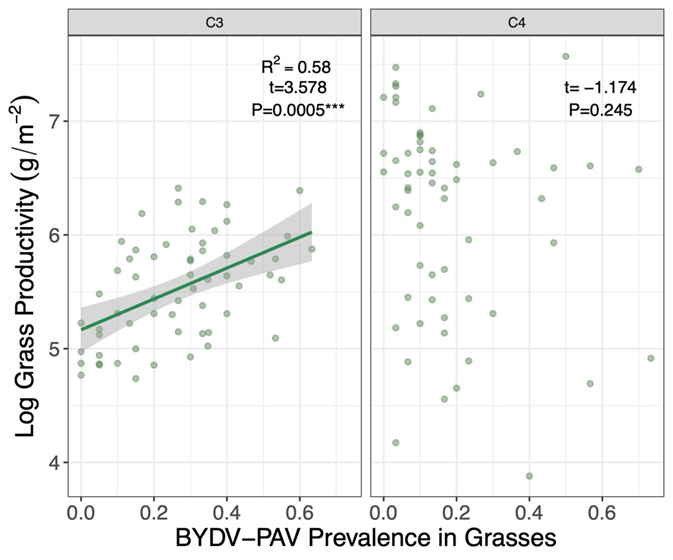
The positive relationship between virus prevalence and C3 grass production may undermine the protective effects of diversity against barley yellow dwarf virus. Several mechanisms could underlie this relationship, including variation in species’ growth rates, species’ net production, and mutualistic pathogen-host interactions. Fast-growing species generally produce more leaf tissue, which can directly increase availability of susceptible leaf tissue for pathogens and create more pathogen transmission opportunities, especially for vector-borne pathogens. Similarly, host net production, the total tissue biomass a species accumulates, can influence pathogen infection dynamics by providing large reservoirs of infected tissue. Lastly, virus-plant interactions may have favourably altered host physiology, contributing to the observed positive relationship between community virus prevalence and net C3 grass production. While plant viruses have been documented to interact mutualistically with their hosts, whether a virus interacts mutualistically or pathogenically can depend on the pathogen’s virulence in the host or on the abiotic stress experienced by the host. However, more work is needed to better understand the conditions under which plant viruses act as pathogens for some hosts and potentially act as mutualists for others.

